Hemmeligheter ved aktiv transport blir synlige
David Coppedge, 20. mars 2024. Oversatt herfra
 Aktiv transport -evnen til å bevege molekyler mot en konsentrasjonsgradient-er en av de viktigste kjennetegnene mellom liv og ikke-liv. Passiv transport, som med osmose, kjenner vi av erfaring: En væske vil naturlig spre seg gjennom en semipermeabel membran fra et område med høy konsentrasjon til en av lav konsentrasjon, inntil konsentrasjonen er utjevnet. Derfor vil bromtabletter i et spa spre seg fra filteret ut i vannet. Det er hvorfor brannrøyk vil lekke inn i et rom gjennom eventuelle sprekker i veggen, men ikke utover. Det vil ta Maxwells demon -lenke, å bekjempe denne naturlige tendensen som følger fra den andre loven om termodynamikk -lenke.
Aktiv transport -evnen til å bevege molekyler mot en konsentrasjonsgradient-er en av de viktigste kjennetegnene mellom liv og ikke-liv. Passiv transport, som med osmose, kjenner vi av erfaring: En væske vil naturlig spre seg gjennom en semipermeabel membran fra et område med høy konsentrasjon til en av lav konsentrasjon, inntil konsentrasjonen er utjevnet. Derfor vil bromtabletter i et spa spre seg fra filteret ut i vannet. Det er hvorfor brannrøyk vil lekke inn i et rom gjennom eventuelle sprekker i veggen, men ikke utover. Det vil ta Maxwells demon -lenke, å bekjempe denne naturlige tendensen som følger fra den andre loven om termodynamikk -lenke.
Livet kan ikke operere etter osmoseprinsippet. En celle med en passiv osmotisk membran vil dø. Celler må aktivt bringe inn eller utvise stoffer, ofte tvinge dem mot en sterk konsentrasjonsgradient. De må opprettholde pH -homeostase uavhengig av forhold utenfor, ofte pumpe inn kationer som Na+, K+, Ca2+og Mg2+eller anioner som klorid CL - selv når interiøret allerede har en mye høyere konsentrasjon enn det ytre. Ved osmose ville denne konsentrasjonen raskt utjevnes og livet ville stoppe. I en virkelig forstand innebærer livet en konstant kamp mot termodynamisk entropi, ved å bruke energi for å bekjempe hva naturkrefter ville gjøre.
Bilde 1. Del av sikkerhetskontroll på flyplass
Unaturlig utvalg
 Biokjemikere har lenge visst om eksistensen av spesialiserte membrankanaler der aktiv transport foregår, og visste at de var svært effektive, men hvordan de opererte var lenge et mysterium. Roderick Mackinnon var en forsker som begynte å finne ut mekanismene for aktiv transport på 1990 -tallet. Han vant Nobelprisen i 2003 for sine oppdagelser om "selektive filtre" innenfor ione-kanaler som lar noen molekyler passere, men ikke andre. Siden den gang har fremskritt innen superoppløselig mikroskopi avslørt detaljer i nesten atomiske skalaer om hva som kan kalles "unaturlig utvalg" i disse kanalene.
Biokjemikere har lenge visst om eksistensen av spesialiserte membrankanaler der aktiv transport foregår, og visste at de var svært effektive, men hvordan de opererte var lenge et mysterium. Roderick Mackinnon var en forsker som begynte å finne ut mekanismene for aktiv transport på 1990 -tallet. Han vant Nobelprisen i 2003 for sine oppdagelser om "selektive filtre" innenfor ione-kanaler som lar noen molekyler passere, men ikke andre. Siden den gang har fremskritt innen superoppløselig mikroskopi avslørt detaljer i nesten atomiske skalaer om hva som kan kalles "unaturlig utvalg" i disse kanalene.
Membrankanaler blir ofte navngitt i henhold til molekylene de transporterer: anion eller kationkanaler, natriumkanaler, kaliumkanaler, kloridkanaler, akvaporiner (vannkanaler) og andre. La oss undersøke den indre virkningen av en kloridkanal, som forskeres kunnskap er oppdatert mot. Vi kan dele spenningen ved å oppdage hvordan det "selektivitetsfilteret" bestemmer hvilke ioner som får lov til å passere. Som en teaser kan du vurdere at selektivitetsfilteret til en kaliumkanal er mye mindre enn bredden på et kaliumion, men det kan transportere 100 millioner ioner per sekund -lenke.
CFTR -klor-kanalen
Forrige måned i PNA -lenke kunngjorde Levring og Chen "strukturell identifikasjon av et selektivitetsfilter i CFTR", en kloridkanal som er ansvarlig for væskehomeostase i epitelvev. Det kalles CFTR (Cystisk Fibrose Transmembran ledningsRegulator) på grunn av den dødelige sykdommen som oppstår når genetisk defekt hindrer passering av kloridioner. På det andre ytterpunktet gjør kolera kanalen for udiskriminerende, noe som fører til diaré som forårsaker dehydrering og død. Nyresykdom kan også være et resultat av mangelfulle CFTR -kanaler. Dette er ikke en del å rote med!
Formen på CFTR ser ut som et buet overflødighetshorn, med en smal innsnevring inni. Legg merke til hvordan forfatterne identifiserer presise aminosyrerester (indikert med en bokstav og et posisjonsnummer) langs kanalbanen som samhandler med kloridionene som går gjennom:
"I denne studien identifiserer vi et kloridbindingssted i de ekstracellulære endene av transmembran-helikser 1, 6 og 8, hvor et dehydatert klorid er koordinert av reststoffene G103, R334, F337, T338 og Y914. Endringer i dette stedet, i samsvar med dets funksjon som et selektivt filter, påvirker ione-selektivitet, konduktans og åpen kanalblokk. Dette selektivitetsfilteret er tilgjengelig fra cytosolen gjennom en stor indre vestibyle og åpner for det ekstracellulære løsningsmidlet gjennom en smal portal. Identifiseringen av et kloridbindingssted på det intra- og ekstracellulære bropunktet, fører til at vi foreslår en komplett konduktans-sti som tillater dehydaterte kloridioner å krysse lipid-vedhengslaget."
Bilde 2. Særegenheter ved celle-membraner (M. Denton)
Diagrammer av interiøret viser et kloridion som lager elektrostatiske kontakt med aminosyrerester på sin reise, som om du kjører en beskyttelseshanske gjennom væpnede vakter som alle sikrer at den har en gyldig tillatelse til å passere. Strukturen “omslutter en kontinuerlig ledning over membranen for klorid for å gjennomtrenge den elektrokjemiske gradienten.” Hvert kloridion er hydrert med en vannkappe, men må fjerne jakken på vei gjennom:
"Hydert klorid kommer inn i den indre vestibylen fra cytosol gjennom en lateral portal mellom TM-er [TransMembrane domener] 4 og 6…. Klorid forblir hydrert i den indre vestibylen og stabiliseres av et positivt elektrostatisk overflatepotensial. Bredden på vestibylen avtar og konvergerer mot et selektivt filter, der bare dehydert klorid kan komme inn. Dehydrert klorid beveger seg inn i dette selektivitetsfilteret, stabilisert ved interaksjoner med G103, R334, F337, T338 og Y914 og rehydrerer ved avkjørsel i epitellumen gjennom en smal sideutgang mellom TMS 1 og 6."
Presisjonsgodkjenning
Hvordan filtrerer kanalen ut andre anioner? Fluor (atomnummer 9) er mindre enn klor (atomnummer 17), så hvorfor glir det ikke gjennom? Forfatterne testet autentiseringsevnen til CFTR ved hjelp av aminosyre-substitusjoner. De bekreftet at fire rester i kanalen utfører en kvalifiseringstest på innkommende anioner når de dehydrerer:
"Som tidligere ble rapportert, og i samsvar med at gjennomsyrende anioner må dehydreres, viser villtype CFTR en lyotropisk permabilitetssekvens, med relativ permabiliteter omvendt relatert til entalpien ved dehydrering ... ved R334a, F337A, T338A, eller Y914F erstatning, ble de relative anion-permabilitetene alle endret, om enn i forskjellige grader, i samsvar med tidligere arbeid."
Figur 5 i artikkelen viser et kloridion som blir inspisert av fire aminosyrerester “COPS” på fire sider. Et useriøst molekyl kommer ikke til å passere! Presisjonen til dette filteret er forbløffende. Hvor mange mutasjon kan systemet tåle uten sammenbrudd? Og hvor mange ulykker bygde dette filteret ved en tilfeldighet i utgangspunktet? Detaljer i følgende sitat vil ikke være på quizen, men for å få en følelse av kompleksiteten som er involvert, kan du se på hvor mange rester som deltar i autentisering av kloridioner når de tester 'beskyttelseshansken':
 "Tidligere mutasjonsarbeid har identifisert et sted med rester, mange er arginin og lysin, som påvirker CFTR-ionelektivitet og/eller konduktans. Kartlegging av disse restene på CFTR -strukturen indikerer at basiske rester, inkludert K95, R104, R117, K190, R248, R303, K335, R352, K370, K1041 og R1048 ... er plassert langs cytosoliske og extracellulære 'vestibyler utsatt for løsningsmiddel. Ulike for restene som direkte koordinerer klorid [selektivitetsfilteret], er funksjonen til disse arginin- og lysinrestene å stabilisere de delvis hydrerte anionene gjennom elektrostatiske interaksjoner og diskriminere mot kationer. -dipol -interaksjoner med klorid.
"Tidligere mutasjonsarbeid har identifisert et sted med rester, mange er arginin og lysin, som påvirker CFTR-ionelektivitet og/eller konduktans. Kartlegging av disse restene på CFTR -strukturen indikerer at basiske rester, inkludert K95, R104, R117, K190, R248, R303, K335, R352, K370, K1041 og R1048 ... er plassert langs cytosoliske og extracellulære 'vestibyler utsatt for løsningsmiddel. Ulike for restene som direkte koordinerer klorid [selektivitetsfilteret], er funksjonen til disse arginin- og lysinrestene å stabilisere de delvis hydrerte anionene gjennom elektrostatiske interaksjoner og diskriminere mot kationer. -dipol -interaksjoner med klorid.
Sidekjedene til Q98 og S341 møter også den cytosoliske vestibylen for å danne anion -dipol -interaksjoner med klorid og bidra til ioneselektivitet. R334, plassert ved porens ekstracellulære munn, spiller en dobbel rolle i å danne selektivitetsfilteret og tiltrekke anioner inn i porene gjennom elektrostatiske interaksjoner. Mange andre funksjonelt viktige rester, inkludert P99, L102, I106, Y109, I336, S1118 og T1134…, samhandler ikke direkte med klorid. I stedet danner de en andre koordineringsfære av [selektivitetsfilteret] som sannsynligvis bidrar til strukturere [selektivitetsfilter] rester med passende geometri for å koordinere klorid."
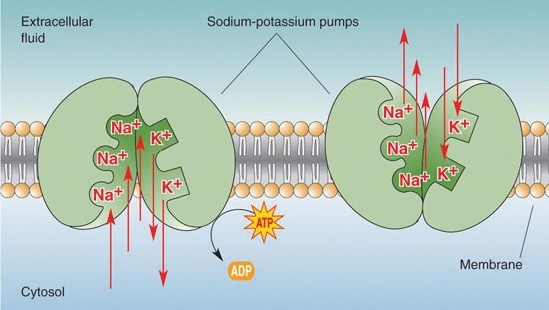
Bilde 3. Ione-kanal (Natrium-Kalium pumper)
Flyplassanalogi
 Tenk på disse andre "viktige restene" som en del av "koordinasjonssfæren" på en flyplass. Hele strukturen tjener formålet med å begrense strømmen av forbipasserende til "selektivitetsfilteret" av røntgenmaskiner som inspiserer passasjerer og bagasjen. Hele overbygningen er nødvendig og må være planlagt med egnet geometri og personell, for å lede passasjerne til inspeksjonsstedet, selv om røntgenmaskinen er like smal som et menneske.
Tenk på disse andre "viktige restene" som en del av "koordinasjonssfæren" på en flyplass. Hele strukturen tjener formålet med å begrense strømmen av forbipasserende til "selektivitetsfilteret" av røntgenmaskiner som inspiserer passasjerer og bagasjen. Hele overbygningen er nødvendig og må være planlagt med egnet geometri og personell, for å lede passasjerne til inspeksjonsstedet, selv om røntgenmaskinen er like smal som et menneske.
TSA -arbeidere på flyplasser kunne aldri skryte av så mye kvalitetskontroll i autentiserings-protokoller. Og menneskelige arbeidere har øyne og sinn til å tenke ut hva de gjør! CFTR -kanalen fungerer automatisk i mørket, ved den delikate "berøringen" av elektrostatiske interaksjoner, innenfor en nøyaktig strukturert smal passasje innenfor koordinasjonsfæren. En kilde sier at CFTR organiserer og behandler millioner kloridioner per sekund! TSA kunne lære noe om effektivitet her, slik mange av oss flypassasjerer kunne attestere.
Når vi snakker om berøring, vil den neste artikkelen min diskutere noen andre kanaler som svarer, når de blir kontaktet-de såkalte mekanosensitive kanalene.
Bilde 4. Aqua-kanaler i gressblad(fra O. Grasso)
Ubrukelig darwinisering
Utviklet CFTR seg? Fordi CFTR -kanalen har noen likheter med andre kloridkanaler som CLC, antar forfatterne overforenkelt at de "unikt utviklet seg fra en familie med aktive transportører", forutsatt at "ikke -relaterte ionekanaler har utviklet seg til å velge og gjennomføre klorid ved bruk av felles kjemiske strategier. En slik fortellings glans er ikke bare ubrukelig, den gir ingen mening. En strategi innebærer framsyn -lenke: å se et behov og designe en løsning. Mens noen hyppige flypassasjerer kan bli fristet til å forestille seg at TSA -strategier virker tankeløse og ikke-styrte, fungerer forseggjorte strukturer som CFTR -kanaler, ekstremt effektivt og nøyaktig med lav toleranse for endring, og ser ingerinørskapte ut. De måtte fungere fra starten av. Uten de nøyaktig plasserte aminosyrerester som allerede er til stede på rett sted innenfor en større koordinasjonsfære, ville det ikke være noen autentisering, og aktiv transport ville stoppe. Alternativet er sykdom og død. Vår enhetlige opplevelse bekrefter at forseggjorte, effektive strategier som fungerer - som bruker ikke-reduserbare komplekse strukturer, med flere koordineringsdeler som støtter funksjonen - alltid er produkter av intelligent design.

Om forfatteren: DAVID COPPEDGE -kredit til David Coppedge (Bilde 5)
David Coppedge er en frilans vitenskapsreporter i Sør-California. Han har vært styremedlem i Illustra Media siden grunnleggelsen og fungerer som deres vitenskapskonsulent. Han jobbet ved NASAs Jet Propulsion Laboratory (JPL) i 14 år, på Cassini-oppdraget til Saturn, til han ble kastet ut i 2011 for å dele materiale om intelligent design, en diskriminerende handling som førte til en nasjonalt publisert rettssak i 2012. Discovery Institute støttet saken hans, men en ensom dommer dømte ham mot ham, uten forklaring. En naturfotograf, friluftsmann og musiker, David har B.S. grader i realfagsutdanning og i fysikk og holder presentasjoner om ID og andre vitenskapelige emner.
Oversettelse, via google oversetter, og bilder ved Asbjørn E. Lund